

進化は枝分かれと交雑で
人類を始めとする生きものは、どのように進化したのか。過去から多くの科学者が探求し続けてきた問いに、化石からゲノムまでを対象に様々なアプローチで研究が進んでいます。この中で私は、目に見える「形」をキーワードにヒトの進化を明らかにしたいと考え、ヒトと同じ霊長類のサルに注目して研究しています。長い時間をかけて進化した霊長類は、何がきっかけで今の多様な姿になったのか。進化の痕跡が隠れているゲノムと、ゲノムと形との関係性を軸に、「種の交わり」を切り口として、難問の解決に繋がる道を探り出したいと考えています。
進化は枝分かれと交雑で
人類を含む霊長類は共通の祖先からヒト(ホモ・サピエンス)やチンパンジー、ゴリラ、オランウータン、ニホンザルなどに分かれて進化してきました。この進化の枝分かれを表現したものが「系統樹」で、枝と枝の間の距離が近ければ近縁種といえます(図1)。系統樹では一本の幹から多くの枝が伸びるように種が分かれており、これはゲノムの変化で身体の形や生理機能が変わり、互いに交配しなくなり別の種になった結果で、「分岐」といいます。一方、ある時点で分かれた種が再び交わって交雑した結果、新しい遺伝子の組み合わせによって多様な表現型が生じることもあります。例えばヒト属では、私たちホモ・サピエンス以外に、北京原人などのホモ・エレクトス、ネアンデルタール人(ホモ・ネアンデルターレンシス)が出現し、ホモ・サピエンス以外はすべて絶滅しました。どの種も別々に進化したと考えられていましたが、古代ゲノム解析の結果から、アフリカ以外のホモ・サピエンス集団とネアンデルタール人の間に遺伝的な交流があったことが示唆されています。つまり、現在のヒトは、絶滅した別の種の特徴を少しだけ受け継いでいる可能性があるのです。
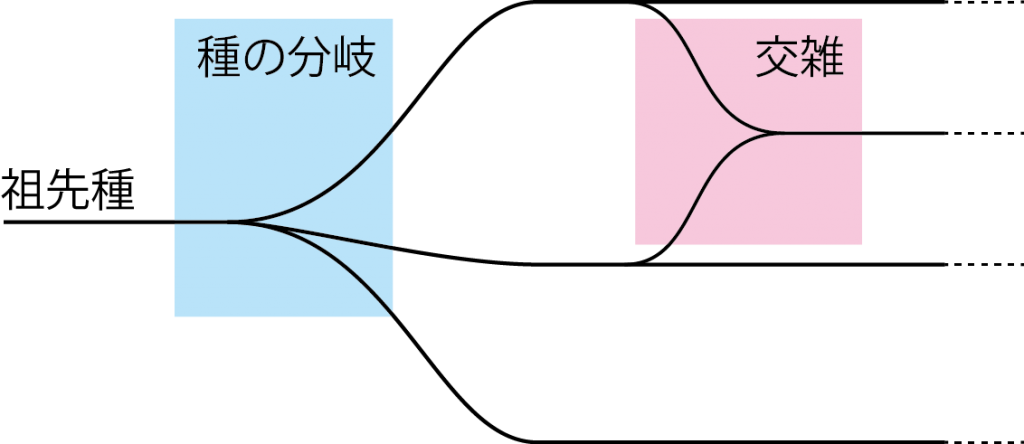
図1
分岐と環境への適応
遺伝子の変化で現れた特徴は、環境に適応すれば後世に残りやすく、適応できなければ残りません。一方、ある特徴が後世に残るか残らないかは、ランダムな偶然の作用によって左右されることもあります。私はこれまでに、ニホンザルなど20種以上を含むマカク属について、形と種の分岐、環境、地域との関係を調べました。マカク属は、熱帯から亜熱帯、温帯まで広い生息地を持つことから、形の違いと環境適応の関係が強く示唆されました。特に「顔」には、目や鼻、口といった環境の変化を感じ取る感覚器官が集まっており、多様な環境へ適応するための変化が大きかったと予測できます。しかし、体の大きさと連動する形の変化が、この関係性を見えにくくしていました。そこで私は、大きさの違いを補正する解析を行い、改めて形と生息環境との関係を調べました。その結果、熱帯に顔の長い種、亜熱帯・温帯に短い種が分布する傾向が見えました。また、分岐関係と照らし合わせたところ、顔の長さの変化は複数回起こっていることが示されたので、地域ごとの変異は偶然ではなく、環境への適応の結果と推察されます(図2)。このように、異なる系統で同じような進化が起こることを「平行進化」といいます。ただ、こうした形の違いがなぜ起こりえたのかについては分かっていません。平行進化の遺伝的メカニズムを明らかにすることが重要で、今後の課題といえます。
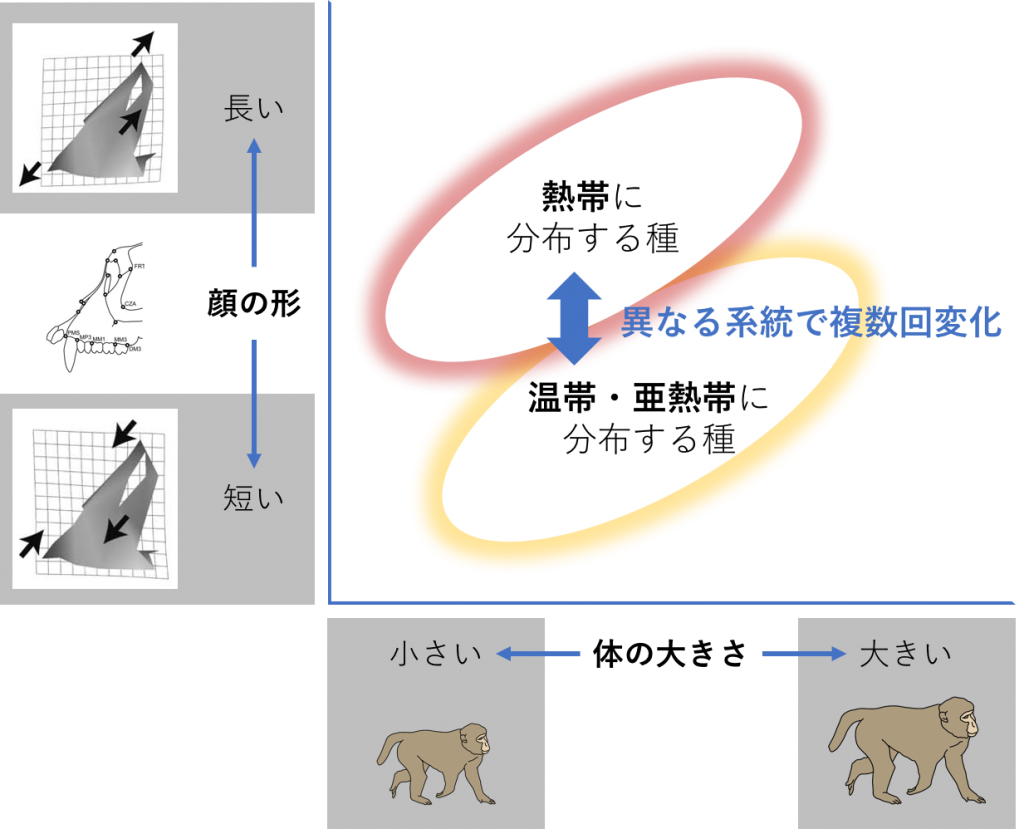
図2
交雑による変化
一方、進化は分岐だけでなく交雑の過程でもあります。ヒトを含めた霊長類において、交雑の遺伝的証拠はいくつも見つかっていますが、交雑がどのように起こって形の進化にどう影響したのかについては、よく分かっていません。人類の進化をひも解こうとしても、ヒト属ではホモ・サピエンス以外が絶滅しているため、交雑の影響を探るための手掛かりがあまりに少ないという制約があります。そこで、ヒト属に比較的近く、種間の交雑が確認できるマカク属に改めて注目しました。マカク属の雑種個体で頭蓋骨の形と交雑度との関係を調べたところ、頭蓋骨の外側の形は両親がもつ特徴の中間型を示す一方で、内側(上顎洞と呼ばれる空洞構造物)は片方の親の形が強く表れていることが分かりました。つまり、外側と内側は異なる遺伝的メカニズムに支配されており、交雑による遺伝子の組み合わせ次第で新しいタイプが出現しうることが考えられます。現在、より詳細に調べるために、頭蓋骨のCT画像データをもとに立体的な3D形態を計測し、ゲノムデータとの関連を調べています(図3)。 また、マカク属では東南アジアのアカゲザルとカニクイザルなどで自然下の交雑が判明しています。近年の技術発展に伴い、サルを捕まえることなく糞からゲノムを解析できるようになりました。マカク属が多く生息する南・東南アジアでのフィールド研究を展開し、ゲノムと形の変異を地域ごとに詳しく調べることで、分岐と交雑、それに伴った形の変化がどのように起こったのか、進化の謎の一端に迫りたいと思います。
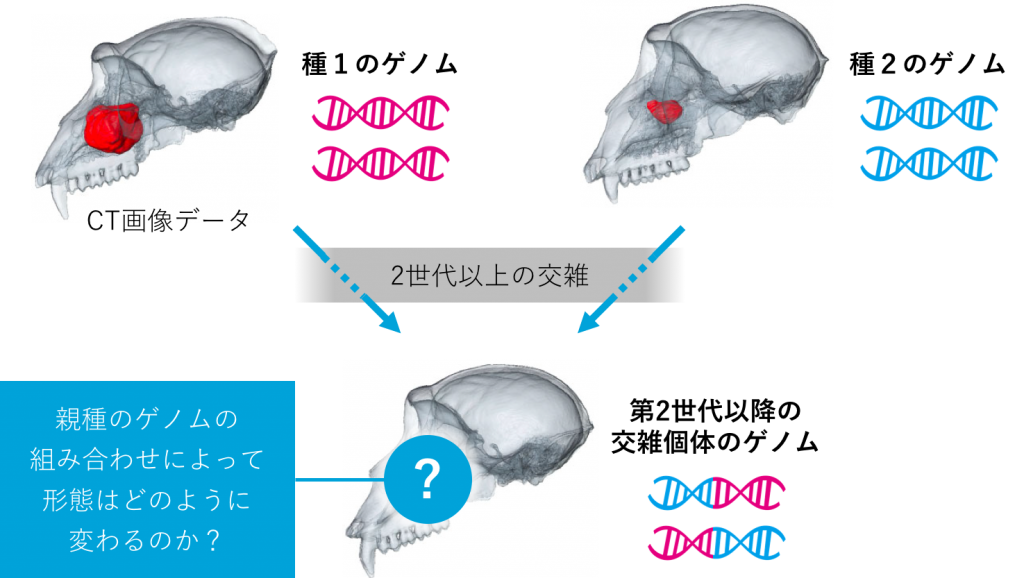
図3
研究室HP
http://www.pri.kyoto-u.ac.jp/shinka/keitai/
主要論文
- Ito T, Kawamoto Y, Hamada Y, Nishimura TD (2015), Maxillary sinus variation in hybrid macaques: implications for the genetic basis of craniofacial pneumatization. Biol J Linn Soc Lond, 115: 333–347. doi:10.1111/bij.12528
- Ito T, Ebbestad JOR, Takai M (2014) Computed tomography examination of the face of Macaca anderssoni (Early Pleistocene, Henan, northern China): Implications for the biogeographic history of Asian macaques. Journal of Human Evolution 72: 64-80.
- Ito T, Nishimura T, Takai M (2014) Ecogeographical and phylogenetic effects on craniofacial variation in macaques. American Journal of Physical Anthropology 154(1):27-41.